Michigan vs. Tasmanian isopod rates of evolution
The Michigan isopod (posted 6/5/2014) with the egg appendage was able to complete two generations per year.
The Tasmanian isopod (posted 6/1/2014) took three years to complete a generation.
Since generation time is a reasonable measure of potential evolutionary rates, there is a six-fold difference in the potential rate of change from the ancestral type between the two groups. Thus it is reasonable to expect the Tasmanian ones have greater similarity to the common ancestor of the two groups. The greater change in the Michigan one was reflected in the flattening of the group, the fusion of abdominal segments of the upper surface of the exoskeleton, and the development of appendages on the egg; these changes do not appear to be evident in the mostly likely ancestral types of crustaceans.
Potential rates of change are just that, potential. Selection may keep a well-adapted set of species characteristics relatively unchanged for a longer time than expected. In such a case, molecular changes may occur in DNA sites not readily affected by selection and thus be a better gauge of time of separation from ancestral types than indicated by unchanged anatomical features.
In ten generations, if x were the numbers of nucleotide changes per generation, the Tasmanian one could have accumulated 10x number of changes and the Michigan one 60x number of changes. Changes to 100% of the genome would take six times longer in the slowest, so more species could be expected to develop in one with a shorter generation time, if rate of potential change were the only operative factor.
SPECIATION CONSEQUENCES OF LONG GENERATION TIME
Speciation, or development of new species, is usually thought to require geographic isolation from the ancestral type. Species are sometimes defined as having reproductive isolation from each other. Geographic isolation can make interbreeding impossible and has been a major factor in evolution. But isopods having a three year generation could have three species going separate evolutionary paths in the same location. I wondered if it may have been a factor in the presence of multiple species of similar isopods in Great Lake in the central highlands of Tasmania. I was not able to determine that although Nicholls listed several species from Great Lake.
Cicadas, some species of which have 17-year life cycles, have developed identifiable differences in some different year-class broods found in the same location. Speciation may have occurred in some invertebrates of the seashore based on different breeding times during the same year by selection differing at the beginning and end of the breeding season, along with higher predation on early life stages by predators focused on the peak at the middle of the breeding season..
In addition to geographic and temporal isolation insects are spectacular in their diversity that is a result in part of rapidly gaining reproductive isolation due to specification of genetalia modifications often referred to as a "lock and key" arrangement. As a result hybridization is prevented once genital differences are sufficient. Insects may also be reproductively isolated by life on different- host plants or animals, or with pheromones, behavior, and microhabitat differences.
WHY DOES LONG GENERATION TIME OCCUR?
I wondered why such long generation time occurred in the Tasmanian isopods. As a result I became sensitive to the causes of long generation time. That followed me long after studying isopods. I had concluded that the rapid input of much nutrients enabled Michigan isopods to specialize for rapid life cycle, with quick growth and the ability to produce many eggs to capitalize on the period of abundant food. At the same time it enabled them to persist in the presence of high levels of predation. So I was familiar with the biology of r- and K- selection before R. H. MacArthur and E. O. Wilson were first to write about it in such publications as their book, The Theory of Island Biogeography (1967). Their elaboration was a valuable contribution to better understanding of the impact of ecology on evolution.
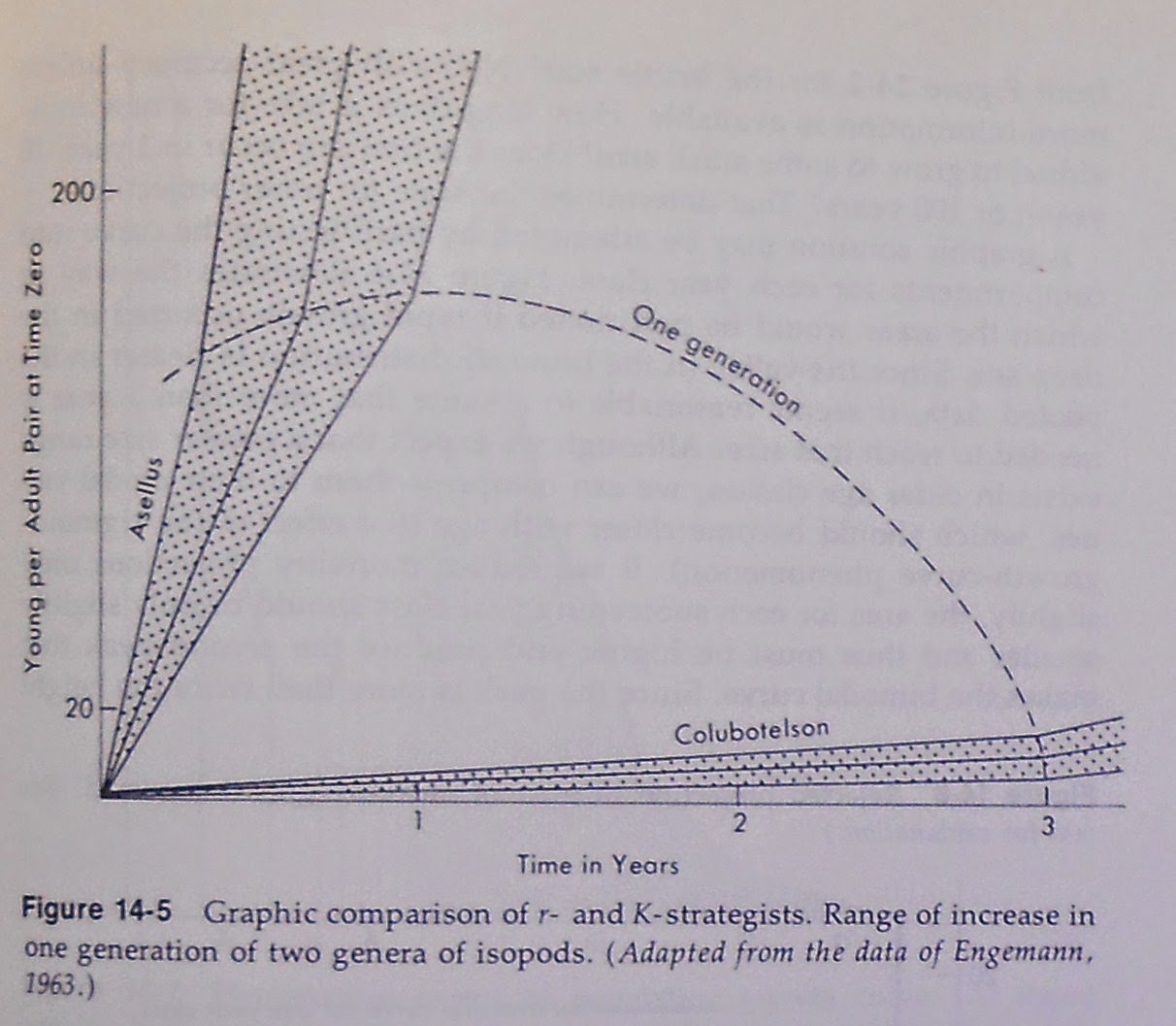
Long life, infrequent and delayed reproduction, low reproductive rates, low food supply, absence of or low predation and/or ways of avoiding predation, and slow development are all associated with K-selected extremes in the biology of a species. Within the same habitat it is possible to have both extremes represented as in the example of the cicada with a dozen or more years needed to complete its life cycle while another heteropteran insect, the aphid, can have many generations per year. Large size usually is accompanied by long generation time, but not always.
WHERE THE ABOVE LED ME
As a result of this interest in longevity, I was prepared in advance to recognize the extreme length of life of the pogonophorans. Their slow evolution and ancestral role were also able to be seen due to that and the knowledge of invertebrates partially acquired by revising Hegner's Invertebrate Zoology text for its 1968 edition. A continuing interest in animal evolution led me to recognize some of the errors infecting it now as shown in the post of May 31, 2013.
Joseph G. Engemann June 9, 2014
No comments:
Post a Comment